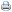Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O trabalho foi realizado na Embrapa-Centro Nacional de Pesquisa de Gado de Leite, no Município de Coronel Pacheco, MG, durante o período de fevereiro a novembro de 2001. Uma área de pastagem de capim-elefante, dividida em piquetes, foi manejada em sistema rotacionado utilizando vacas mestiças Holandês x Zebu em lactação (5 animais/ha). O cronograma de pastejo utilizado durante o período experimental constituiu-se em três dias de ocupação/piquete e 30 dias de descanso. A altura do resíduo pós-pastejo foi, em média, de 1,0 m.
Aplicaram-se a lanço 200 kg de N e de K2O e 50 kg de P2O5 por hectare (1.000 kg/ha da fórmula 20-5-20), divididos em três aplicações iguais durante o período de novembro de 2000 a março de 2001 (novembro, janeiro e março).
As variáveis climáticas observadas durante os períodos de avaliação, obtidas em posto meteorológico localizado a aproximadamente 1 km de distância do local do experimento, são apresentadas na Tabela 1.
O estudo de morfogênese foi realizado segundo o delineamento de blocos casualizados, com seis repetições. Foram realizadas avaliações durante os meses de fevereiro/março, abril/maio, julho/agosto e outubro de 2001.
Na avaliação das taxas de aparecimento, alongamento (Mazzanti et al., 1994) e senescência (Andrade, 2001) de folhas, foram identificadas três touceiras representativas nos seis piquetes e, em cada touceira, foram identificados dois perfilhos (um basilar e outro aéreo). Cada repetição foi constituída pelo valor médio dos três perfilhos de cada tipo em cada piquete, totalizando seis repetições.
A lâmina foliar foi medida em seu comprimento até sua completa expansão, ou seja, até o aparecimento da lígula. O comprimento da lâmina emergente foi medido do seu ápice até a lígula da última folha expandida, até que sua lígula se tornasse visível. As observações foram feitas a cada três ou quatro dias. Foi definido como data de aparecimento foliar o dia em que se observou a exposição da lígula (Andrade, 2001) e a data de morte quando todo seu comprimento se apresentava necrosado (Vine, 1983; Gomide & Gomide, 2000). Tais observações e registros, realizados durante o período de descanso, iniciaram-se após a saída dos animais do piquete pastejado e se repetiram em cada ciclo de pastejo, até a próxima utilização do piquete.
Foram avaliadas as taxas de aparecimento, alongamento e senescência foliar por perfilho. Foi possível caracterizar também o número total de folhas e de folhas vivas por perfilho. A vida útil das folhas foi estimada tomando-se por base valores de suas taxas de aparecimento e número de folhas por perfilho (Chapman & Lemaire, 1993). A taxa de alongamento foliar (cm/dia/perfilho) foi obtida dividindo a diferença entre o comprimento total final de lâminas foliares e comprimento total inicial, pelo número de dias envolvidos na avaliação (período de descanso da pastagem). A taxa de aparecimento de folhas (folhas/dia/perfilho) foi calculada pela divisão do número de folhas totalmente expandidas (lígula exposta) surgidas por perfilho pelo número de dias envolvidos. O inverso da taxa de aparecimento de folhas estimou o intervalo de tempo, em dias, para aparecimento de folhas no perfilho (filocrono). A taxa de senescência foliar (cm/dia/perfilho) foi calculada dividindo o comprimento final total do tecido senescente pelo número de dias envolvidos.
Na estimativa da densidade de perfilhos por m2 foi feita a contagem do número de touceiras em uma área de 49 m2. Nesta mesma área, três touceiras representativas tiveram seus perfilhos aéreos e basilares contados para a determinação do número de perfilhos. A densidade de perfilhos por m2 foi obtida com a divisão do produto destas duas contagens pela área em que as touceiras foram contadas (49 m2).
A produção de biomassa foliar foi obtida por duas técnicas: 1) Pastejo simulado, feito em três touceiras representativas de cada piquete, das quais a primeira representou as de porte baixo, a segunda, as de porte médio e a terceira, as de porte alto. Todas as folhas de cada touceira foram colhidas manualmente, e em seguida pesadas e secadas em estufa a 65ºC para determinação da matéria seca. O peso seco médio das folhas das três touceiras multiplicado pelo número de touceiras contadas em 49 m2 foi usado para a estimativa da disponibilidade de forragem; 2) A partir das estimativas da densidade de perfilhos totais e das taxas de alongamento de folhas, ou seja, amostras de lâminas foliares, emergentes e expandidas, após medição do comprimento, foram secadas em estufa a 65ºC e pesadas, a fim de se estabelecer o índice gravimétrico, isto é, o fator de conversão entre comprimento e peso seco de lâmina foliar (mm de lâmina foliar/perfilho transformado em mg de matéria seca de lâmina foliar/perfilho) (Mazzanti et al., 1994; Gomide, 2001). Esse procedimento permitiu estimativas das taxas de crescimento e senescência de folhas por perfilho. Assim, a produção foi estimada pela contribuição do crescimento de folhas (F), sendo F = taxa de alongamento foliar x população de perfilhos x índice gravimétrico x dias de crescimento.
Os dados obtidos foram submetidos à análise de variância, para verificar os efeitos de época do ano nas características morfogênicas e estruturais de perfilhos basilares e aéreos, e as médias comparadas pelo teste de Tukey a 5% de probabilidade. Na avaliação do efeito de tipos de perfilho (aéreos e basilares), considerou-se os valores médios das épocas do ano, que foram comparadas pelo teste t a 5% de probabilidade.
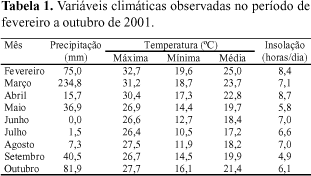
A taxa de aparecimento de folhas foi maior em perfilhos basilares que em perfilhos aéreos (Tabela 2). Os intervalos de tempo médios para o aparecimento de folhas foram de 6,3 e 7,5 dias/folha para perfilhos basilares e aéreos, respectivamente. Gomide (1997) relatou variação de 2,4 a 10,0 dias/folha, dependendo da espécie, do manejo e da época do ano. Entretanto, os valores observados neste trabalho se aproximam dos obtidos por Andrade (2001) para a cultivar Napier (7,1 dias/folha) e por Almeida et al. (2000) para a cultivar Mott (6,5 dias/folha).
A taxa de aparecimento de folhas variou com a época do ano e os maiores valores foram constatados durante o período fevereiro/março e os menores, durante julho/agosto, independentemente do tipo de perfilho (Tabela 2). Os intervalos médios de aparecimento de folhas, considerando perfilhos aéreos e basilares, foram de 4,9, 6,7, 12,8 e 6,8 dias, respectivamente, durante o período de fevereiro/março, abril/maio, julho/agosto e outubro. O filocrono varia conforme a espécie forrageira, mas, para um mesmo genótipo, responde à época do ano em decorrência das condições de luz, temperatura e umidade no solo (Wilhelm & McMaster, 1995). O maior período para aparecimento de folhas durante julho/agosto evidencia a situação desfavorável das plantas ao serem submetidas à baixa precipitação pluvial e diminuição da temperatura e do fotoperíodo.
A taxa de alongamento de folhas variou conforme o tipo de perfilho, sendo o maior valor médio obtido para o basilar em relação ao aéreo (Tabela 2).
Em relação às épocas do ano, os dois tipos de perfilhos apresentaram comportamento semelhante, com maior taxa de alongamento de folhas durante o verão, quando as condições climáticas foram mais favoráveis ao crescimento da gramínea. Foi observada queda acentuada durante o inverno, quando a temperatura e a precipitação pluvial decresceram acentuadamente (Tabela 1). Andrade (2001) constatou redução de 45% na taxa de alongamento de folhas do capim-elefante não irrigado em relação ao irrigado. No presente trabalho, foi observado decréscimo na taxa de alongamento mais acentuado que o encontrado por Andrade (2001). No campo, as variáveis ambientais atuam simultaneamente no crescimento da gramínea, intensificando os efeitos do clima nas características morfogênicas. Assim, os efeitos causados pela seca podem estar associados a outros fatores tais como, temperaturas supra-ótimas ou redução na disponibilidade de nutrientes, que normalmente se verifica em condições de limitação hídrica (Nabinger & Pontes, 2001).
A taxa de senescência foliar não variou com a época do ano em perfilhos basilares. Já em perfilhos aéreos, esta taxa foi maior em julho/agosto e menor em fevereiro/março. Quanto ao tipo de perfilho, foi observado maior valor médio em perfilhos basilares do que em perfilhos aéreos (Tabela 2). Andrade (2001) encontrou taxas de senescência de folhas de capim-elefante de 4,96 e 1,46 cm/dia/perfilho, respectivamente, em perfilhos basilares e aéreos. Segundo esse autor, a taxa de senescência foi baixa até a idade de 52 dias, aumentando linearmente após este período. Isto explica a acentuada diferença entre os valores observados neste trabalho, em que as avaliações se restringiram ao período de descanso da pastagem (30 dias), e os obtidos por Andrade (2001).
As estimativas da vida útil das folhas foram de 27, 35, 31 e 30 dias, respectivamente, em fevereiro/março, abril/maio, julho/agosto e outubro, e de 29 e 28 dias em perfilhos aéreos e basilares, respectivamente. Com base no critério da vida útil das folhas para definição da freqüência de utilização do pasto, a fim de minimizar as perdas por senescência (Parsons & Penning, 1988), deduz-se que o período de descanso de 30 dias, usado neste trabalho, foi adequado, uma vez que se aproximou dos valores estimados.
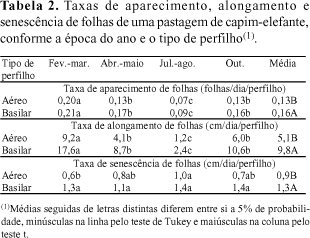
Folhas mais curtas foram observadas em julho/agosto, independentemente do tipo de perfilho (Tabela 3). Em perfilhos aéreos foi observada menor variação do tamanho da folha que em perfilhos basilares. O tamanho final da folha é determinado pela relação taxa de alongamento/taxa de aparecimento de folhas. A taxa de aparecimento de folhas é inversamente correlacionada com o tamanho final da folha (Nabinger & Pontes, 2001). Aparentemente, poderia se esperar folhas menores em fevereiro/março, quando as taxas de aparecimento foram maiores. Como o comprimento das folhas está diretamente relacionado à sua taxa de alongamento, as elevadas taxas de alongamento, observadas nesta época, contribuíram para obtenção de folhas de maior tamanho.
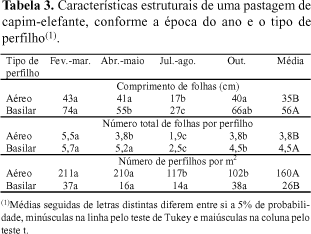
O número total de folhas por perfilho atingiu valor máximo de 5,7 em perfilhos basilares e 5,5 em perfilhos aéreos, ao final dos 30 dias do período de descanso (Tabela 3). Em ambos os tipos de perfilhos, maiores números de folhas foram observados em fevereiro/março e menores em julho/agosto. Quanto ao número de folhas vivas expandidas por perfilho, observaram-se 4,9 folhas nos perfilhos basilares e 4,0 folhas nos aéreos. A estabilização do número de folhas vivas por perfilho é sugerida como critério na determinação do momento de corte da planta e do período de descanso do pastejo rotativo (Fulkerson & Slack, 1995; Gomide, 2001). É provável que o número de folhas vivas por perfilho ainda não tivesse se estabilizado ao final do período de descanso da pastagem, em virtude do curto período de avaliação. Andrade (2001) observou estabilização do número de folhas vivas em perfilhos de capim-elefante, apenas aos 51 dias de idade. A adoção de variáveis isoladas para determinação do manejo pode levar a conclusões equivocadas (Lemaire & Agnusdei, 1999). Em pastagens formadas com gramíneas de alongamento precoce do colmo, como o capim-elefante, o manejo deve ser orientado, também, pela relação folha/colmo, por sua relevância quanto ao valor nutritivo e consumo de forragem.
O número de perfilhos variou conforme a época do ano, apenas nos do tipo aéreo. Maiores valores foram observados em fevereiro/março e abril/maio e menores, em julho/agosto e outubro (Tabela 3). O número de perfilhos basilares não variou com a época do ano e foi menor que o de perfilhos aéreos. Os valores extremos variaram de 102 a 211 perfilhos aéreos e de 14 a 38 perfilhos basilares. O mais intenso perfilhamento aéreo do capim-elefante é uma característica desejável, considerando que esse tipo de perfilho tem menor tendência para elevar o meristema apical, maior proporção de folhas e maior densidade de forragem por cm de altura da planta em comparação com os perfilhos basilares (Hillesheim & Corsi, 1990).
As taxas de acúmulo e a produção total de biomassa foliar foram maiores em fevereiro/março do que nas demais épocas (Tabela 4). Isso reflete, em grande parte, as maiores taxas de alongamento foliar, característica morfogênica de alta correlação com o rendimento forrageiro (Horst et al., 1978).
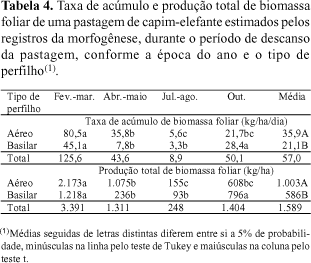
Os perfilhos aéreos apresentaram maiores acúmulos médios de biomassa do que os basilares. A contribuição dos perfilhos aéreos para a produção total de biomassa foliar foi de 63%, considerando os valores médios das épocas do ano. Esses resultados revelam que as maiores taxas de alongamento e aparecimento de folhas, observadas em perfilhos basilares, não compensaram a menor quantidade desse tipo de perfilho. Por essa razão, os perfilhos aéreos foram os que mais contribuíram para a produção de biomassa de folhas.
O acúmulo de biomassa foliar durante o período de descanso da pastagem foi estimado pela morfogênese, tomando-se por base as taxas de alongamento de folhas e as densidades de perfilhos, e pela técnica do pastejo simulado. Não foi observada diferença entre as estimativas de disponibilidade de folhas em fevereiro/março, abril/maio e julho/agosto. Já em outubro, o valor obtido por meio das variáveis morfogênicas foi maior que aquele estimado pelo pastejo simulado (Tabela 5).

Os rendimentos forrageiros obtidos neste trabalho são semelhantes aos observados por Aroeira et al. (1999) e Deresz (2001) em trabalhos que estimaram a disponibilidade de biomassa de capim-elefante por meio do pastejo simulado. Sob o ponto de vista de nutrição animal, as técnicas aqui relatadas apresentam a vantagem de estimar valores próximos da biomassa potencialmente consumível por animais em pastejo, considerando a preferência dos ruminantes em consumirem folhas (Chacon et al., 1978).
1. A época do ano influencia as variáveis morfogênicas e estruturais e o acúmulo de biomassa foliar no pasto de capim-elefante.
2. A maior parte da biomassa foliar do capim-elefante, manejado em sistema de pastejo rotativo com 30 dias de descanso, é resultado do crescimento de perfilhos aéreos.
3. A produção de biomassa foliar pode ser obtida a partir das estimativas da densidade de perfilhos e suas taxas de alongamento de folhas.
ALMEIDA, E. X.; MARASCHIN, G. E.; HARTHMANN, O. E. L.; RIBEIRO FILHO, H. M. N.; SETELICH, E. A. Oferta de forragem de capim-elefante anão "Mott" e a dinâmica da pastagem. Revista Brasileira de Zootecnia,Viçosa, MG, v. 29, n. 5, p. 1281-1287, 2000. www.scielo.br
ANDRADE, A. C. Morfogênese, análise de crescimento e composição bromatológica do capim-elefante (Pennisetum purpureum Schum. cv. Napier) adubado e irrigado sob pastejo. 2001. 81 f. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa, MG, 2001.
AROEIRA, L. J. M.; LOPES, F. C. F.; DEREZ, F.; VERNEQUE, R. S.; MALDONADO VASQUEZ, H.; MATOS, L. L.; VITTORI, A. Pasture availability and dry matter intake of lacting crossbred cows grazing elephantgrass (Pennisetum purpureum Schum.). Animal Feed Science and Technology, Amsterdam, v. 78, p. 313-324, 1999.
CARVALHO, P. C. F.; RIBEIRO FILHO, H. M. N.; POLI, C. H. E. C.; MORAES, A.; DELAGARDE, R. Importância da estrutura da pastagem na ingestão e seleção de dietas pelo animal em pastejo. In: SOCIEDADE BRASILEIRA DE ZOOTECNIA (Piracicaba, SP). A produção animal na visão dos brasileiros. Piracicaba: Fealq, 2001. p. 853-871.
CHACON, E.; STOBBS, T. H.; DALE, M. B. Influence of sward characteristics on grazing behaviour and growth of Hereford steers grazing tropical grass pasture. Australian Journal of Agricultural Research, Collingwood, v. 29, n. 1, p. 89-102, 1978.
CHAPMAN, D. F.; LEMAIRE, G. Morphogenetic and structural determinants of plant regrowth after defoliation. In: INTERNATIONAL GRASSLAND CONGRESS, 17., 1993, Palmerston North. Proceedings Palmerston North: SIR Publishing, 1993. p. 95-104.
DERESZ, F. Influência do período de descanso da pastagem de capim-elefante na produção de leite de vacas mestiças Holandês x Zebu. Revista Brasileira de Zootecnia, Viçosa, MG, v. 30, n. 2, p. 461-469, 2001. www.scielo.br
FULKERSON, W. J.; SLACK, K. Leaf number as a criterion for determining defoliation time for Lolium perene - II: effect of defoliation frequency and height. Grass and Forage Science, Oxford, v. 50, n. 1, p. 16-29, 1995.
GOMIDE, C. A. M. Características morfofisiológicas associadas ao manejo do capim-mombaça (Panicum maximum Jacq.). 2001. 107 f. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa, MG, 2001.
GOMIDE, C. A. M.; GOMIDE, J. A. Morfogênese de cultivares de Panicum maximum Jacq. Revista Brasileira de Zootecnia, Viçosa, MG, v. 29, n. 2, p. 341-348, 2000. www.scielo.br
GOMIDE, J. A. Morfogênese e análise de crescimento de gramíneas tropicais. In: SIMPÓSIO INTERNACIONAL SOBRE PRODUÇÃO ANIMAL EM PASTEJO, 1997, Viçosa, MG. Proceedings... Viçosa, MG: UFV, 1997. p. 411-429.
GRANT, S. A.; MARRIOT, C. A. Detailed studies of grazed sward-techniques and conclusions. Journal of Agricultural Science, Cambridge, Inglaterra, v. 122, n. 1, p. 1-6, 1994.
HILLESHEIM, A.; CORSI, M. Capim-elefante sob pastejo: fatores que afetam as perdas e utilização de matéria seca. Pesquisa Agropecuária Brasileira,Brasília, v. 25, n. 9, p. 1233-1246, set. 1990.
HORST, G. L.; NELSON, C. J.; ASAY, K. H. Relationship of leaf elongation to forage yield of tall fescue genotypes. Crop Science, Madison, v. 18, n. 5, p. 715-719, 1978.
LEMAIRE, G.; AGNUSDEI, M. Leaf tissue turn-over and efficiency of herbage utilisation. In: GRASSLAND ECOPHYSIOLOGY AND GRAZING ECOLOGY, 1999, Curitiba. Proceedings... Curitiba: UFPR, 1999. p. 165-186.
MAZZANTI, A.; LEMAIRE, G.; GASTAL, F. Effect of nitrogen fertilization upon herbage production of a tall fescue sward continuously grazed by sheep - 1: herbage growth dynamics. Grass and Forage Science, Oxford, v. 49, n. 3, p. 111-120, 1994.
NABINGER, C.; PONTES, L. S. Morfogênese de plantas forrageiras e estrutura do pasto. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, Piracicaba, 2001. Anais... Piracicaba: Fealq, 2001. p. 755-770.
PARSONS, A. J.; PENNING, P. D. The effect of duration of regrowth on photosynthesis, leaf death and the average rate of growth in a rotational grazed sward. Grass and Forage Science, Oxford, v. 43, n. 1, p. 15-27, 1988.
PEDREIRA, C. G. S.; MELLO, A. C. L.; OTANI, L. O processo de produção de forragem em pastagem. In: SOCIEDADE BRASILEIRA DE ZOOTECNIA (Piracicaba, SP). A produção animal na visão dos brasileiros. Piracicaba: Fealq, 2001. p. 772-807.
VINE, D. A. Sward structure changes within a perennial ryegrass sward: leaf appearance and death. Grass and Forage Science, Oxford, v. 38, p. 231-242, 1983.
WILHELM, W. W.; McMASTER, G. S. Importance of the phyllochron in studying development and growth in grasses. Crop Science, Madison, v. 35, n. 1, p. 1-3, 1995.
Domingos Sávio Campos PaciulloI; Fermino DereszII; Luiz Januário Magalhães AroeiraII; Mirton José Frota MorenzIII; Rui da Silva VernequeII - rsverneq[arroba]cnpgl.embrapa.br
IEmbrapa-Centro Nacional de Pesquisa de Gado de Leite (CNPGL), Rua Eugênio do Nascimento, 610, Dom Bosco, CEP 36038-330 Juiz de Fora, MG. Bolsista do CNPq. E-mail: dscp[arroba]terra.com.br IIEmbrapa-CNPGL. E-mail: deresz[arroba]cnpgl.embrapa.br, laroeira[arroba]cnpgl.embrapa.br, rsverneq[arroba]cnpgl.embrapa.br IIIUniversidade Estadual do Norte Fluminense, Centro de Ciências e Tecnologias Agropecuárias, Av. Alberto Lamego, s/no, CEP 28015-620 Campos dos Goytacazes, RJ. E-mail: morenz[arroba]uenf.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|